
Remember me



Group 2 innate lymphoid cells (ILC2s), known for their role in defending against parasites and regulating tissue homeostasis through cytokine secretion, were found to have a new function. Research in Cell shows that, when cultivated in a lab, human ILC2s can target and destroy tumor cells in a living organism using various cell death mechanisms.
Innate lymphoid cells (ILCs) represent a critical component of the innate immune system. These cells are categorized into 5 canonical subsets based on their cytokine production profiles and functional roles.1 ILC1s and natural killer (NK) cells produce interferon-gamma and cytotoxic molecules for tumor surveillance and defense against viral infections. ILC2s play a pivotal role in the immune response to parasitic infections through their secretion of interleukins such as IL-5 and IL-13. ILC3s are particularly important in defense against bacterial pathogens, especially in the gut, and they contribute to tissue repair and homeostasis by producing cytokines like IL-17 and IL-22. Lymphoid tissue inducer cells are critical for the formation of lymph nodes. Dysregulation of ILC function can contribute to pathological conditions.2
Of these ILC subsets ILC2s are probably the best studied one. Beyond their role in immune responses they contribute to restoring tissue integrity following injury and play a role in regulating lipid metabolism and thermogenesis, especially in adipose tissues.3
Like other ILC subsets, ILC2s are highly plastic.3 They can transition from a state of maintaining tissue integrity and supporting organ function to an activated state where they actively participate in inflammatory responses. This transition is influenced by the local cytokine environment and allows ILC2s to rapidly respond and adapt to various environmental cues, including cytokines, allergens, and infectious agents. Such responsiveness enables these cells to shift their functions to meet the specific demands of the immune response.
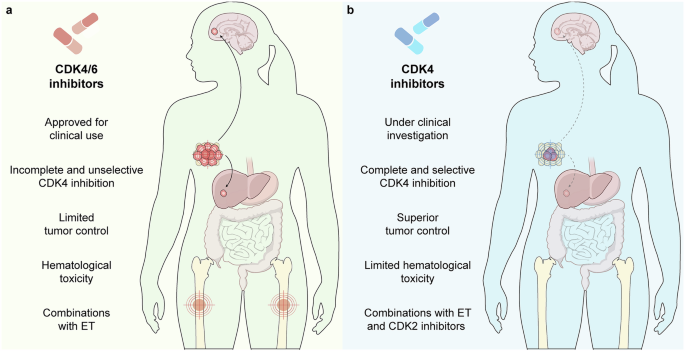
Recent studies have highlighted the multifaceted roles of ILC2s in cancer immunity.3 Although ILC2s have been observed to facilitate tumor progression in mouse models of hepatocellular carcinoma and colorectal cancer (CRC),4 they also appear to indirectly contribute to the rejection of tumors in mouse models of melanoma, as well as pancreatic cancer and CRC, by enhancing the responses of cytotoxic T cells and eosinophils (Fig. 1a).5,6
Fig. 1: Roles of ILC2s in cancer immunity.
a In mice, ILC2s indirectly contribute to antitumor immunity in models of melanoma, pancreas cancer and CRC (further reviewed in3). b Li et al. demonstrate how human ILC2s, in contrast to mouse ILC2s, can be activated to directly lyse tumor cells in AML as well as glioblastoma, pancreas and lung cancer. Mechanistically, CD155-induced DNAM-1 expression on ILC2s leads to GZMB and perforin (PRF1) expression and thereby tumor apoptosis and pyroptosis. Prolonged exposure of ILC2s to CD155 leads to downregulation of DNAM-1 and reduced cytotoxicity, which is prevented by the anti-cancer drug Trametinib. Created with BioRender.com.
A paper published recently in Cell highlights a new role for ILC2s with implications for cancer immunotherapy.7 Previous research showed that human ILC2s can produce the cytotoxic molecule Granzyme B (GZMB) when stimulated by alarmins, which are primarily produced by epithelial cells.8 Li and colleagues show that human ILC2s, when expanded in vitro, not only continue to express GZMB but also develop the ability to produce another cytolytic molecule, perforin (Fig. 1b). This endows them with innate immune cytolytic abilities similar to NK cells, specifically against tumor cells. This finding is particularly striking when compared to mouse ILC2s, which lack this cytolytic function, underscoring the value of cross-species research. Mechanistically, human ILC2s trigger Caspase-3-mediated apoptosis in tumor cells via perforin and GZMB release. Additionally, GZMB is capable of cleaving gasdermin E (GSDME) in GSDME-positive tumor cells, leading to pyroptosis.
A key focus of the study is the role of the DNAM-1 (CD226) receptor on human ILC2s. This receptor, crucial in antitumor immunity, interacts with its ligands CD112 and CD155, which are highly expressed on tumor cells. The interaction enhances GZMB production in ILC2s, leading to the induction of pyroptosis and/or apoptosis in acute myeloid leukemia (AML) cells. IL-15, which was shown in another study to induce GZMB and perforin in ILC3s, may amplify this. For DNAM-1, this mechanism includes the phosphorylation and inactivation of the inhibitor FOXO1, boosting GZMB production in ILC2s. The authors also show a role for NKG2D, which interacts with its ligands MICA/MICB/ULBP1/2/5/6 on tumor cells, in inducing GZMB production in ILC2s. However, the downstream signaling of this interaction was not as extensively explored as for DNAM-1.
However, the study also reveals a challenge in AML patients as they show lower expression of DNAM-1 and GZMB in ILC2s compared to healthy donors. As an explanation for this observation, prolonged exposure of AML blasts to CD155 leads to internalization and degradation of DNAM-1 in ILC2s. This could impair the recognition and immune clearance of AML. The study suggests that blocking CD155 might prevent tumor escape from various DNAM-1-positive immune cells, including ILC2s. Indeed, the mitogen-activated protein kinase kinase (MEK) inhibitor trametinib, which reduces expression of CD155 on AML blasts, prevented the downregulation of DNAM-1 expression on ILC2s.
The study raises the prospect of using human ILC2s in cancer immunotherapy. Given the challenges associated with CAR-T cell therapy, including toxicities and the high relapse rate in AML, the study proposes the use of expanded ILC2s as allogeneic, off-the-shelf therapies. These could be used immediately upon AML patient remission, potentially avoiding relapse and the delays associated with autologous CAR-T cell preparation. Cytotoxic ILC2s demonstrated activity in vitro not only against AML cells, but also against cell lines derived from pancreatic, brain, and lung tumors. This suggests the potential for a wider use of human ILC2s in immunotherapy for various solid cancers.
There is much to uncover about cytolytic ILC2s, including what regulates their perforin expression to deliver GZMB in target cells. GZMB+ ILC2s have been successfully generated in vitro using the OP9 mouse stromal cell line, which expresses the Notch ligand Delta Like 1, along with cytokines IL-2, IL-7, and IL-15. However, the natural counterparts of in vitro generated cytolytic ILC2s, expressing both GZMB and perforin, remain elusive in living organisms. It is possible that ILC2s in tumor tissues express both GZMB and perforin, thereby possessing cytolytic potential but this has yet to be determined. The role of cytolytic ILC2s in immune responses, especially against non-tumor pathogens like viruses, is yet to be clarified. This exploration faces challenges due to their apparent non-existence in mice. Nevertheless, the ubiquity of CD226 ligands in tissues suggests that cytolytic ILC2s could be more prevalent than currently believed, and their presence in inflamed human tissues could offer insights into their functions.
In conclusion, the study demonstrates that ILC2s can acquire cytolytic functions similar to ILC1s9 and ILC3s10 further underscoring the highly plastic nature of these cells. Moreover, new insights are provided into the antitumor properties of human ILC2s, not observed in mouse ILC2s, and indicate their potential as a cytolytic immune effector in cancer. This opens up new avenues for treating both hematological and solid tumors by harnessing the unique capabilities of ILC2s.
Comments (0)