Plants are sophisticated bio-factories for providing humans with energy, fibers, drugs, etc. The demand for plant-based products is rising due to the increase of human population, exceeding the natural capacity of plants, which is particularly true for many bioactive specialized metabolites produced within certain medicinal plants [1]. Catharanthus roseus is an important natural source owing to its exclusive potential to synthesis over one hundred terpenoid indole alkaloids (TIAs) which are extensively used in the treatment of various chronic diseases [2]. Among these TIAs, vinblastine and vincristine, which bind to tubulin and inhibit microtubule assembly during expansion of cancer cells [3], were only found in C. roseus so far [4]. The chief limitation in the production of TIAs is that critical metabolites, such as vinblastine and vincristine, were produced at very low levels [5]. As though de novo microbial biosynthesis of vindoline and catharanthine in engineered yeast, following with in vitro chemical coupling to vinblastine, has been achieved [6]. The cost for synthesis of TIAs was high due to their complicated structure, and the extraction from plants was still the main strategy for commercial production of these metabolites [7]. The very low content of these valuable compounds in C. roseus made them extremely expensive [8]. Furthermore, vincristine and vinblastine were listed as being drugs with a shortage in 2019–2020 by the US Food and Drug Administration, there is growing awareness of the importance to promote their production [9].
Considering TIAs' low yield in C. roseus and high cost, great research efforts were made to enhance their production through various biotechnology-based strategies. As alkaloids could act as defense compounds in plants under stress [10], elicitations both biotically and abiotically are one of the most promising strategies to alter the production of secondary metabolites in C. roseus [11]. The TIAs biosynthetic pathways have been clearly characterized and a 38-step pathway sequentially presented in at least three distinct cell types in leaves was revealed, as well as the potential regulation mechanism [12]. Many transcription factors (TFs) were found to regulate the expression of genes encoding different enzymes in the TIAs pathway. The octadecanoid responsive Catharanthus AP2/ERF (ORCA) TFs such as ORCA2, ORCA3, ORCA4 and ORCA5 were one of the most recognized TFs involved in the regulation of TIA biosynthesis [[13], [14], [15], [16]]. In addition, the regulatory function of many other TFs belonging to WRKY superfamily, MYB superfamily, G-box binding TFs, zinc finger TFs, and basic helix-loop-helix TFs were characterized as well [17]. Various “push and pull” strategies aiming to regulating the metabolic flux, individually or in combination modifying biosynthetic genes and related transcriptional regulators, have been used to boost TIA production [11]. While transcriptional factors ORCA3, BIS1, and MYC2 were shown to regulate genes of different rate-limited enzymes in the TIA pathway, the co-overexpression of these three TFs had a limited effect on downstream TIAs [18]. Another most recent findings on a new TIA regulator CrMYC1 indicated that the overexpression of CrMYC1 in hairy root line significantly improved the accumulation of ajmalicine and catharanthine [19], which provided insight into the role of this understudied TF in TIA pathway regulation and might hint further research objectives. Overall, the regulatory network of TIA accumulation has not been fully elucidated.
Nucleus is important organellar for being the regulatory hub of gene expression in response to various environmental stress, where nuclear receptors triggered dynamic signaling pathways [20]. Multi-omics technologies provided rich data for better understanding of stress response within plants; however, while transcriptomic techniques are characterized by high sensitivity sequencer that allow for the detection of low-expressed transcripts such as nuclear transcriptional factors [21], detections of low-abundant proteins are still limited as their signals are largely masked by abundant proteins [22]. In order to investigate the crucial biological functions of nuclear proteins by proteomic methods, enriching nuclear fractions from the whole cell has been a useful tool for achievement of high nuclear proteome coverage as well as precise quantification [23]. Yin and Komatsu [24] enriched nuclear fractions to study upstream events controlling the regulation of flooding-responsive proteins in soybean, where the decrease of chromatin structure-, pre-ribosome biogenesis-, and mRNA processing-related proteins suppressed protein translation. Barua et al. [25] studied nuclear proteins and phosphoproteins from chickpea subjected to gradual dehydration and revealed predominance of differential phosphorylation of proteins related to splicing, protein degradation, regulation of flowering time, and circadian. Lamelas et al. [26] isolated and quantified nuclear proteins in heat-stressed seedlings of Pinus radiata, which discovered that epigenetic mechanisms played a key role in stress tolerance and priming mechanisms. Ayash et al. [27] identified >100 low abundant transcription factors and transcriptional co-activators from nuclear proteome of Arabidopsis thaliana treated with elicitors of pattern triggered immunity, as well as several hundred dual targeted protein, which suggested protein import into the nucleus in plant immunity. As though plant nuclear proteome contained information related to physiological function under various environmental stress, the nuclear proteome of C. roseus remained poorly studied.
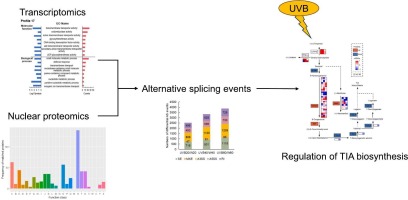
Various environmental factors, such as light [28], drought [29], temperature [30], etc., have been proved to have regulatory effect of TIA biosynthesis. Ultraviolet (UV) radiation is a form of non-ionizing radiation that originally emitted by the sun and is classified into three groups based on their wavelength. Previous researches suggested that UVB induced TIA biosynthesis in C. roseus seedlings [31], suspension cells [32], and hairy roots [33]. To investigate UV-response mechanisms in C. roseus, proteome and phospho-proteome comparisons between radiated plants and untreated plants were analyzed, which identified dynamic protein phosphorylation induced by increase of ATP, followed with increase of protein related to protein modification, calcium-dependent signaling, and biosynthesis of monoterpene precursor [[34], [35]]. As these findings preliminarily suggested that the UV-B-associated signal transduction pathway were involved in the regulation of TIA pathway, molecular regulation at different intervals during prolonged UVB was not clear.
Therefore, in order to decipher more comprehensive molecular characteristics in C. roseus under UVB radiation, the transcriptome data at different intervals of UVB radiation were obtained; Furthermore, nuclear fractions of C. roseus were enriched and identified by mass spectrometry. Correlations between the transcriptomic and the proteomic data were analyzed; and conjoint analysis was performed. The results would help elucidate the regulatory mechanisms in C. roseus in response to UVB radiation, which could be useful for engineering high-TIA-yielding system.









留言 (0)