
記住我

Gastroduodenal ulcer, also known as peptic ulcers, are characterized by the formation of ulcers within the mucosal lining of the stomach or duodenum. Clinical manifestations typically encompass upper abdominal discomfort, accompanied by symptoms such as nausea, vomiting, and dyspepsia (Malfertheiner et al., 2009). Frequent etiological factors encompass Helicobacter pylori infection, the use of nonsteroidal anti-inflammatory drugs (NSAIDs), aspirin, and genetic predisposition (Huang et al., 2002; Graham, 2014; Miftahussurur and Yamaoka, 2015). Notwithstanding our deepening comprehension of the pathophysiological underpinnings of Gastroduodenal ulcer and the progress in therapeutic modalities, the persistent issues of extended treatment duration, post-treatment monitoring, and the specter of recurrence continue to pose substantial challenges. These challenges not only result in financial burdens on patients and their families but also exert a profound impact on patients’ quality of life and occupational productivity (Lanas and Chan, 2017). Furthermore, Gastroduodenal ulcer can give rise to complications, notably perforations, with potentially grave consequences for patients’ well-being (de Boer, 1997). Hence, there is an urgent demand for the exploration of additional therapeutic modalities for Gastroduodenal ulcer with the aim of ameliorating the socioeconomic strain on both families and society.
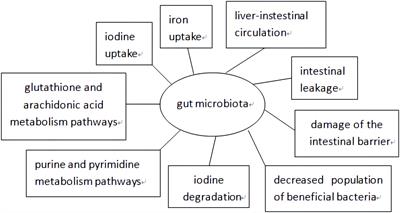
The gut microbiota constitutes an intricate and diversified microbial consortium inhabiting the human gastrointestinal tract. It encompasses a wide array of microorganisms, encompassing bacteria, fungi, viruses, and parasites. Notably, bacteria represent the predominant constituents among them (Sender et al., 2016). The gut microbiota assumes a pivotal role in the human organism. It is involved in the synthesis and secretion of diverse bioactive substances, including vitamins, enzymes, and antibiotics, that confer numerous advantages to human physiology. Furthermore, the gut microbiota actively participates in the processes of digestion, the maintenance of intestinal health, and the preservation of microbial diversity and equilibrium within the gut. These functions are of paramount importance for overall health (Thursby and Juge, 2017). Furthermore, the gut microbiota exerts a pivotal influence on the maturation and modulation of the human immune system, thereby actively contributing to the preservation of immune homeostasis (Belkaid and Hand, 2014). The correlation between the gut microbiota and Gastroduodenal ulcer is an ongoing subject of research. Studies have indicated disparities in the gut microbiota composition between Gastroduodenal ulcer patients and healthy individuals (Chen X. et al., 2018). For instance, patients with Gastroduodenal ulcer may exhibit an overabundance of pathogenic bacteria, such as Helicobacter pylori, in their gut microbiota. Moreover, the disturbance of gut microbiota equilibrium may be associated with Gastroduodenal ulcer development (Schulz et al., 2018).
Due to the potential influence of uncontrolled confounding variables, traditional research methodologies often face limitations in elucidating the precise association between gut microbiota and Gastroduodenal ulcer. Consequently, the establishment of a causal link between gut microbiota and Gastroduodenal ulcer remains inconclusively characterized. Mendelian randomization (MR) serves as a viable approach to assess the presence of a causal relationship between the exposure and the outcome. This is achieved by employing single-nucleotide polymorphisms (SNPs) closely linked to relevant risk factors as instrumental variables (IVs) (Emdin et al., 2017). The reliability of the causal relationship within this methodology stems from the fact that the random allocation of alleles during embryonic meiosis remains largely impervious to the influence of most confounding variables (Zhang et al., 2023). To address the current void in Mendelian randomization (MR) analyses concerning the causal nexus between gut microbiota and Gastroduodenal ulcer, we undertook a comprehensive genome-wide association study (GWAS) followed by a two-sample Mendelian randomization (MR) investigation. This research endeavors to furnish a more profound comprehension of the influence exerted by gut microbiota on Gastroduodenal ulcer and to proffer robust scientific substantiation conducive to the prevention and management of Gastroduodenal ulcer through gut microbiota modulation.
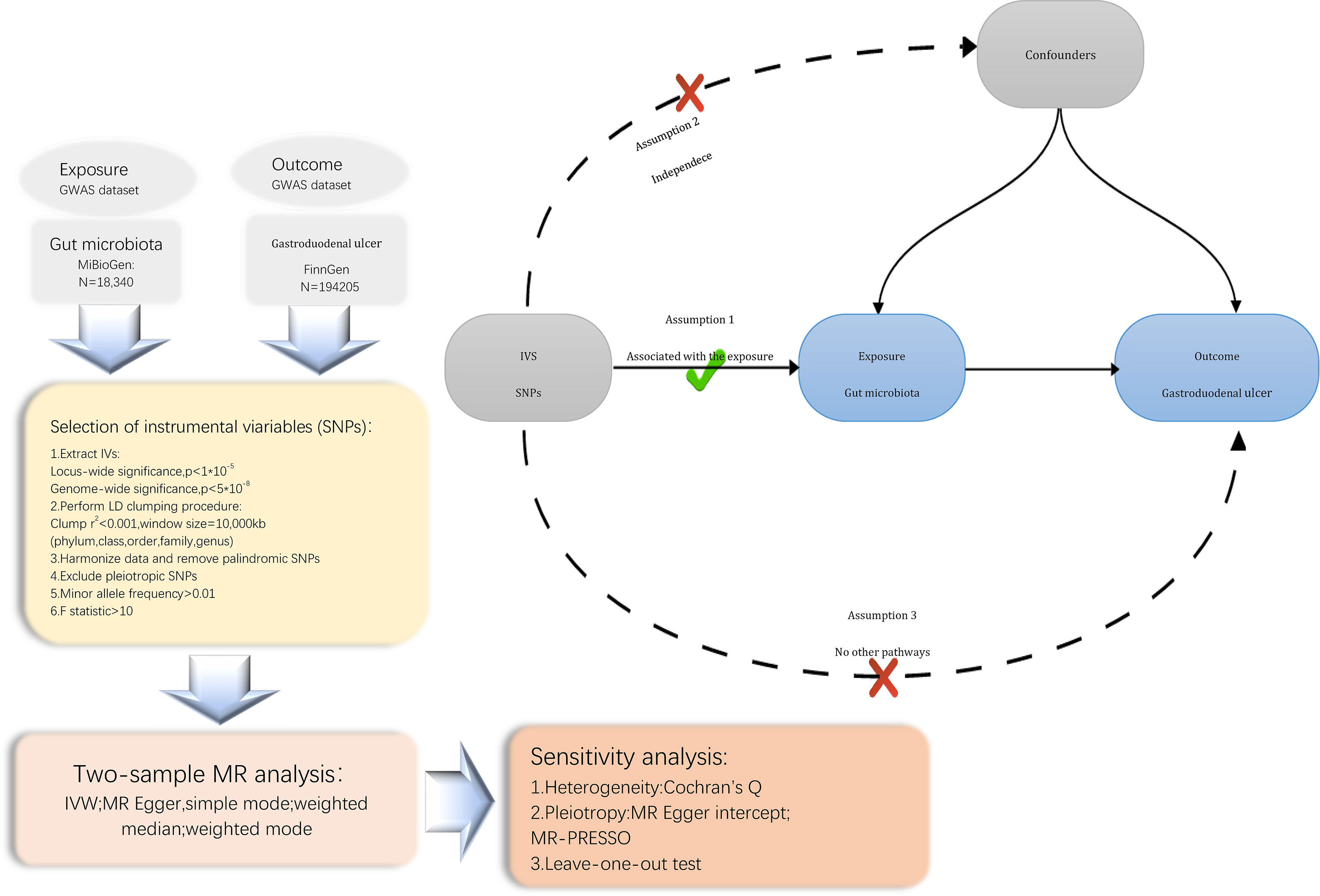
2 Methods2.1 Study design and data sourcesWe conducted a Mendelian randomization (MR) study to explore the causal relationship between the gut microbiota and Gastroduodenal ulcer. The schematic diagram of our study process is shown in Figure 1. In summary, we extracted data from summary statistics of genome-wide association studies (GWAS) to identify genetic variations associated with the exposure, which were subsequently used as instrumental variables (IVs). We performed a sequential two-sample MR analysis employing five distinct MR methodologies. Finally, a comprehensive set of sensitivity analysis metrics, including tests for heterogeneity, pleiotropy, and leave-one-out analysis, were applied to assess significant associations.
Figure 1 Flowchart of the present MR study and major assumptions. MR, Mendelian randomization; GWAS, genome-wide association study; SNPs, single nucleotide polymorphisms; IVW, inverse-variance weighted; LD, linkage disequilibrium; MR-PRESSO, MR pleiotropy residual sum and outlier.
Summary-level genomic data of the gut microbiota were acquired from the MiBioGen study (data from https://mibiogen.gcc.rug.nl/) (Kurilshikov et al., 2021).This study represented the largest and most diverse genome-wide meta-analysis of the gut microbiota to date, encompassing genome-wide genotyping data and 16S fecal microbiota profiles from 24 cohorts, comprising a total of 18,340 individuals. The majority of participants in the study were of European descent (N=13,266). Profiling of microbial composition was achieved through targeted sequencing of the V4, V3-V4, and V1-V2 regions of the 16S rRNA gene. Subsequently, taxonomic classification was performed utilizing direct taxonomic binning. Following the processing of 16S microbiome data, a total of 211 taxa were identified, encompassing 131 genera, 35 families, 20 orders, 16 classes, and 9 phyla. Comprehensive information regarding the microbiota dataset can be found in the original investigation (Kurilshikov et al., 2021).
The GWAS summary data on Gastroduodenal ulcer (finn-b-K11_GASTRODUOULC) were obtained from the FinGen, including 9216 cases of Gastroduodenal ulcer and 320387 controls (Kurki et al., 2023). To enhance the reliability of our findings, we carried out an extensive search in the “ieu open gwas project” focusing on data related to Gastroduodenal ulcer. After careful screening, we selected the dataset with the largest sample size, which not only had a large sample size but also contained detailed information about Gastroduodenal ulcer. Using this large sample dataset, we can analyze and study the relevant features and risk factors of Gastroduodenal ulcer more accurately. Thus, our results will be more persuasive and will provide stronger evidence for research and practices in relevant fields (data from https://gwas.mrcieu.ac.uk/).
2.2 Instrumental variable selectionTo ensure the accuracy and validity of our conclusions regarding the causal relationship between gut microbiota and Gastroduodenal ulcer, we implemented a series of quality control procedures to filter instrumental variables (IVs). Firstly, we selected single-nucleotide polymorphisms (SNPs) with significant associations to the gut microbiome as IVs. SNPs were chosen based on two distinct thresholds. In order to obtain a comprehensive overview and enhance the explained phenotypic variability, we included a set of SNPs with locus-wide significance levels below 1×10-5 as IVs. Additionally, for secondary analysis, another set of SNPs with genome-wide significance (p<5×10-8) were selected as IVs, but we did not find enough sample size in our experiments. Secondly, to ensure the independence of the selected IVs and minimize the impact of linkage disequilibrium that violates the random allele assignment, we configured the clumping procedure with parameters set to r2<0.001 and kb=10,000kb. Thirdly, If exposure-related SNPs were not identified in the outcome genome-wide association study (GWAS) results, proxy SNPs highly correlated with the target variant (r2>0.8) were identified through the SNiPA website (Arnold et al., 2015). However, it’s important to note that such a scenario did not occur in our analysis. Fourthly, SNPs with palindromic properties and incompatible alleles were disqualified from the Mendelian Randomization (MR) analysis. Fifthly, in order to satisfy the second key assumption of MR (independence from confounders), we conducted a manual inspection and exclusion of SNPs significantly associated (p<5×105) with potential confounding factors using the PhenoScanner GWAS database (Staley et al., 2016; Kamat et al., 2019). SNPs such as rs166849 and rs6494306 were eliminated because they were associated with past smoking and type 2 diabetes. In addition, SNPs rs2952251 were associated with anxiety, past smoking, and mood swings, SNPs rs62532512 were associated with past smoking, mood swings, and misery, and SNP rs17708276 were associated with worry, tension, and misery, and all SNPs rs2952251, rs62532512, and rs17708276 were also deleted. Sixthly, a minimum minor allele frequency threshold of 0.01 was enforced. Lastly, to mitigate weak instrumental bias, the F-statistic was computed for each SNP, and any SNPs with F-statistics below 10 were discarded (Burgess and Thompson, 2011). The F-statistic is expressed as R2 (n-k-1)/k (1-R2), with n representing the sample size, k denoting the number of IVs, and R2 signifying the variance explained by the IVs.
2.3 Effect size estimateWe conducted a two-sample Mendelian randomization (MR) analysis to explore the causal relationship between gut microbiome features and the risk of Gastroduodenal ulcer.When multiple IVs were involved in a gut microbiota feature, we adopted the inverse-variance weighted (IVW) test as the primary analytical approach, complemented by other methodologies, including MR-Egger, simple mode, weighted median, and weighted mode (Burgess et al., 2013). To comprehensively assess the influence of the gut microbiome on Gastroduodenal ulcer, the meta-analysis technique known as IVW converted the outcome effects of IVs on exposure effects into a weighted regression model with an intercept constrained to zero. In the absence of horizontal pleiotropy, IVW yielded unbiased estimates by mitigating the influence of confounding variables (Holmes et al., 2017). It is noteworthy that the MR-Egger method may be susceptible to the influence of outlier genetic variables, potentially leading to incorrect estimations. However, even when all selected IVs are invalid, the MR-Egger approach can still produce unbiased estimates (Bowden et al., 2016b). The simple mode offers robustness against pleiotropy effects, although it may be less statistically powerful than IVW (Milne et al., 2017). The weighted median method, when at least 50% of data from valid instruments are available, is capable of providing precise and reliable effect estimates (Bowden et al., 2016a). In situations involving genetic variables that violate the pleiotropy assumption, the weighted mode method can be adapted (Hartwig et al., 2017).
2.4 Sensitivity analysisTo assess the potential impact of heterogeneity and pleiotropy among instrumental variables (IVs) on MR results, a comprehensive set of sensitivity analyses was undertaken to ascertain the robustness of our significant findings. Heterogeneity among the selected genetic instruments was quantified using Cochran’s Q test and visualized through funnel plots. Furthermore, we probed for potential horizontal pleiotropic effects of the included IVs, employing both the MR Egger intercept and the Mendelian randomization pleiotropy residual sum and outlier (MR-PRESSO) global test. Concurrently, we performed a leave-one-out sensitivity analysis to validate the precision and robustness of causal effect estimates, ensuring that our MR estimates were not unduly influenced by highly influential SNPs. In addition, the MR Steiger directionality test was employed to infer the causal direction (Hemani et al., 2017). Credible causal links were identified when the variance explained by the IVs on the exposure exceeded that on the outcome. All statistical analyses in our investigation, encompassing both MR and sensitivity analyses, were executed using the R packages “TwoSampleMR” and “MRPRESSO” within the publicly available R software (version 4.3.1).
3 Results3.1 Instrumental variable selectionIn our study, we commenced by choosing 211 bacterial taxa as the subjects of investigation. To guarantee the adherence of instrumental variables (IVs) to the established criteria, we conducted a rigorous screening process to eliminate instrumental variables that exhibited significant associations with the study objectives. In order to satisfy the second critical assumption of Mendelian Randomization (MR), which pertains to the independence of confounding factors, we further utilized the PhenoScanner GWAS database for a meticulous manual examination. This allowed us to identify and subsequently exclude instrumental variables that displayed significant associations with potential confounding factors. During this process, we carefully identified instrumental variables significantly linked to confounding factors and duly removed these variables to ensure the precision of our research outcomes. These procedures are essential in ensuring the independence of the instrumental variables we employed, allowing us to effectively infer causal relationships. Subsequently, the remaining data underwent re-analysis utilizing the aforementioned methodologies. The results of the analysis indicated that when employing the Inverse Variance Weighting (IVW) method as the primary analytical approach, the p-value associated with Rikenellaceae exceeded 0.05. As a result, Rikenellaceae and all the included Single Nucleotide Polymorphisms (SNPs) were removed. During the final screening phase, we rigorously selected 156 Single Nucleotide Polymorphisms (SNPs) as instrumental variables (see Supplementary Table S1). These instrumental variables underwent meticulous filtration to guarantee their effectiveness and reliability within the context of our study. It is noteworthy that all instrumental variables exhibited F-values exceeding 10, signifying their robust predictive capacity in explaining variables. Importantly, this observation underscores that our instrumental variables are not weak, and they can be effectively employed to address endogeneity issues (see Table 1). These outcomes bolster our confidence in the validity of our research findings and furnish robust support for subsequent analyses.
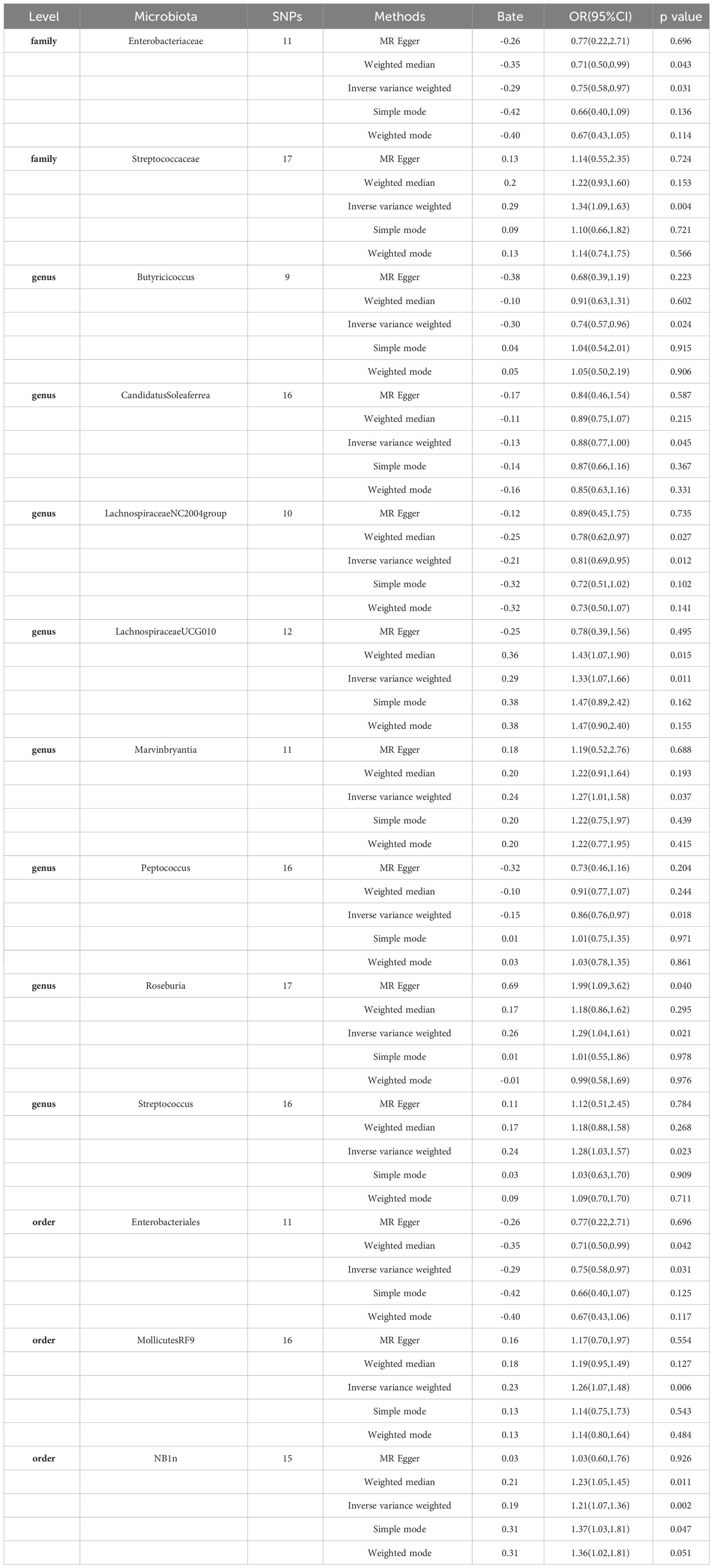
Table 1 MR estimates for the association between gut microbiota and Gastroduodenal ulcer (p < 1 × 10−5).
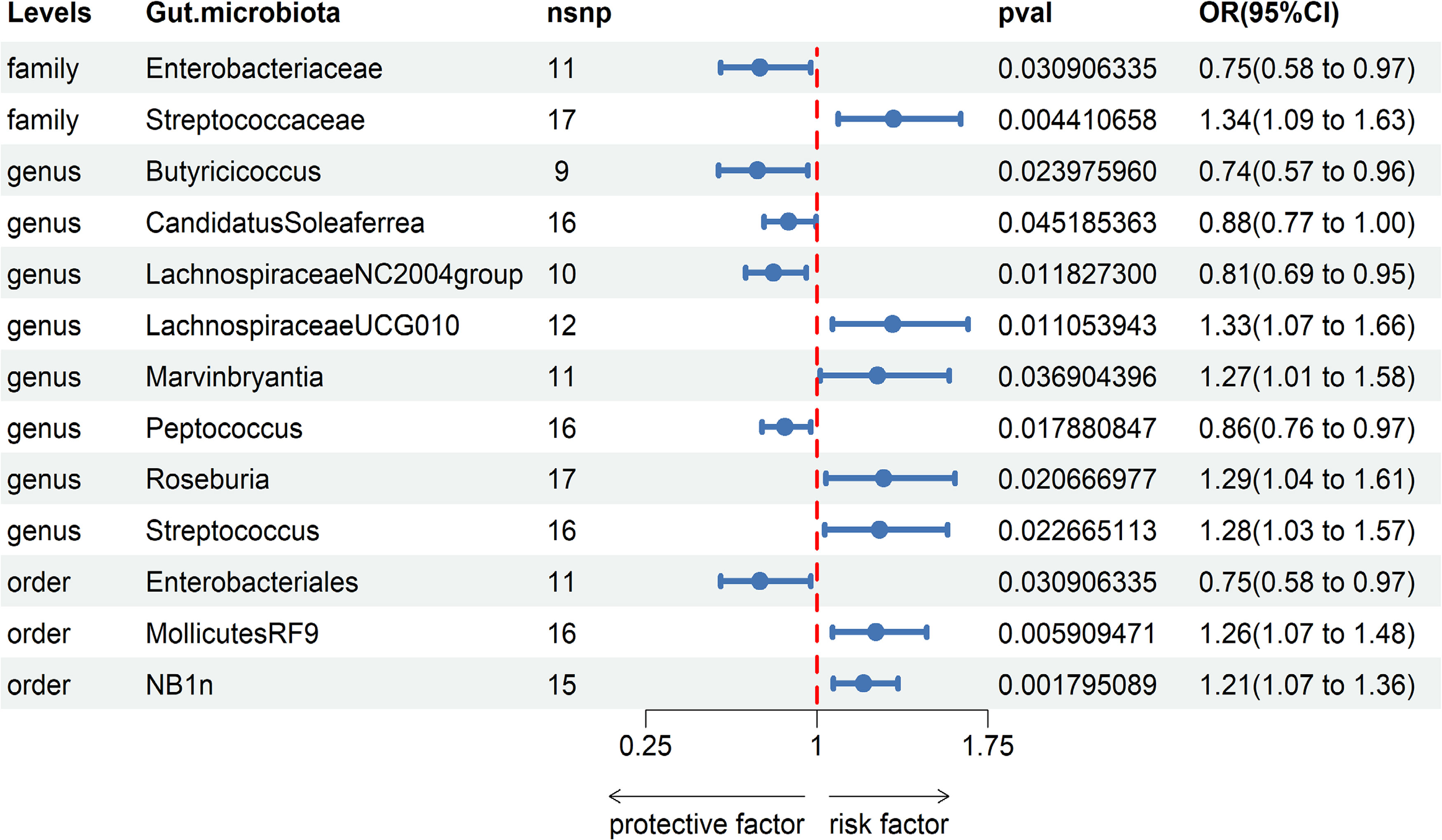
3.2 Causal impact of gut microbiota on gastroduodenal ulcerBased on our research findings, we have identified causal relationships between 13 bacterial genera and the risk of Gastroduodenal ulcer. Notably, several bacterial taxa with high predicted abundance exhibited significant correlations with the risk of Gastroduodenal ulcer.
Specifically, a higher abundance of Enterobacteriaceae (OR: 0.75, 95% CI: 0.58-0.97, p=0.031)was associated with a reduced risk of Gastroduodenal ulcer. Similarly, increased abundances of Butyricicoccus (OR: 0.74, 95% CI: 0.57-0.96, p=0.024), Candidatus Soleaferrea (OR: 0.88, 95% CI: 0.77-1.00, p=0.045), Lachnospiraceae NC2004 group (OR: 0.81, 95% CI: 0.69-0.95, p=0.012), Peptococcus (OR: 0.86, 95% CI: 0.76-0.97, p=0.018), and Enterobacteriales (OR: 0.75, 95% CI: 0.58-0.97, p=0.031) were associated with a decreased risk of Gastroduodenal ulcer (see Figure 2, Table 1).
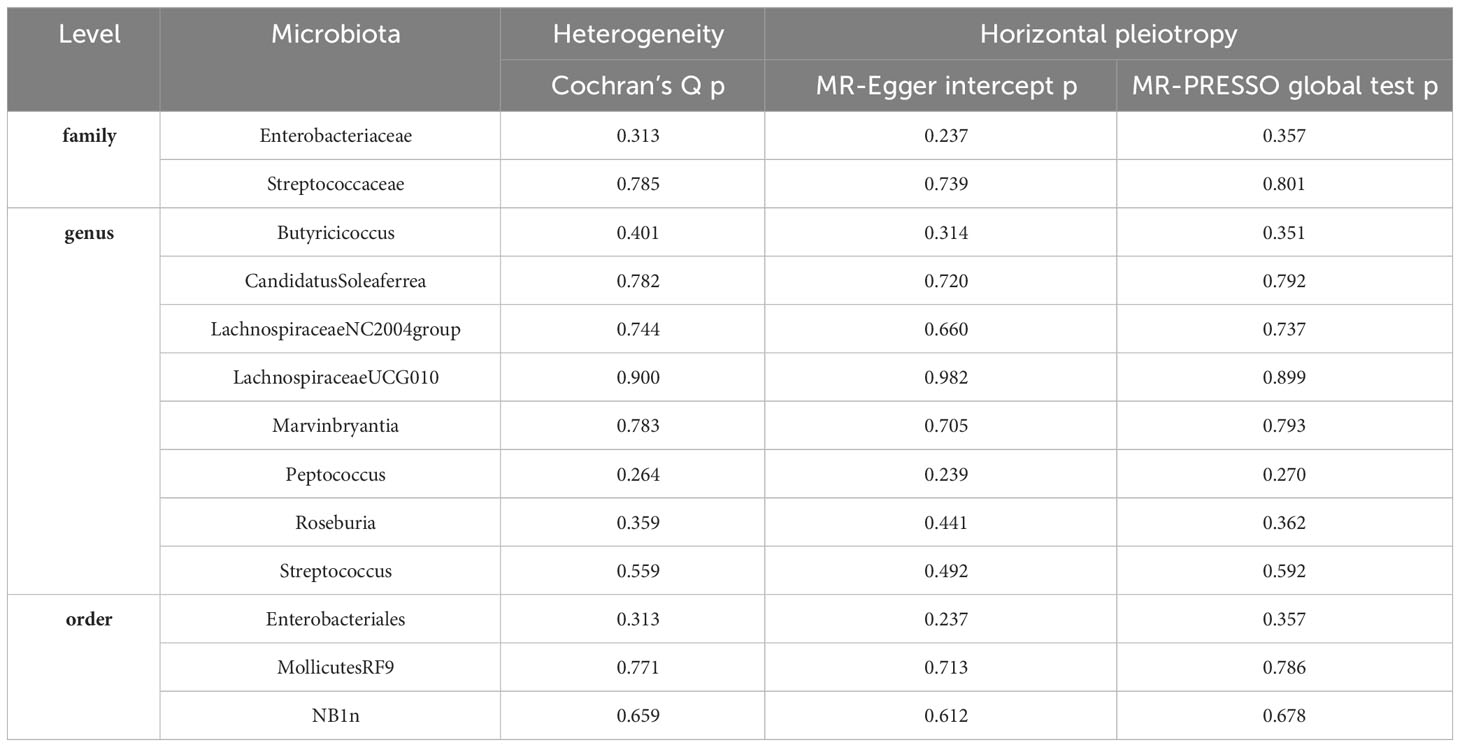
Table 2 Evaluation of heterogeneity and directional pleiotropy using different methods.
In contrast, higher abundances of Streptococcaceae (OR: 1.34, 95% CI: 1.09-1.83, p=0.004), Lachnospiraceae UCG010 (OR: 1.33, 95% CI: 1.07-1.66, p=0.011), Marvinbryantia (OR: 1.27, 95% CI: 1.01-1.58, p=0.037), Roseburia (OR: 1.29, 95% CI: 1.04-1.61, p=0.021), Streptococcus (OR: 1.28, 95% CI: 1.03-1.57, p=0.023), Mollicutes RF9 (OR: 1.26, 95% CI: 1.07-1.48, p=0.006), and NB1n (OR: 1.21, 95% CI: 1.07-1.36, p=0.002) were associated with an elevated risk of Gastroduodenal ulcer. These findings suggest that increased abundances of these gut microbiota may be linked to an increased risk of Gastroduodenal ulcer (see Figure 2, Table 1).
Figure 2 Associations of genetically predicted Gastroduodenal ulcer with sepsis risk using IVW method SNPs, single nucleotide polymorphisms; OR, odds ratio; CI, confidence interval.
3.3 Sensitivity analysisOur analysis of the relationship between gut microbiota and Gastroduodenal ulcer has revealed a total of 13 causal associations. Initially, we employed Cochran’s Q test to assess the heterogeneity of all instrumental variables (IVs). The results indicated no significant heterogeneity (P>0.05), signifying consistent effects of the selected instrumental variables (IVs) across various studies (refer to Table 2).
To further validate the instrumental variables (IVs), we performed MR-Egger intercept tests and MR-PRESSO tests. Encouragingly, all p-values exceeded 0.05, indicating the absence of horizontal pleiotropy and the detection of outliers by MR-PRESSO (refer to Table 2). This absence of outliers further bolsters our confidence in the instrumental variables (IVs). These sensitivity analyses confirm that the selected instrumental variables (IVs) exhibit good heterogeneity and lack horizontal pleiotropy, effectively assisting in addressing endogeneity concerns.
The scatter plot illustrates the relationship between distinct gut microbiota and the incidence of Gastroduodenal ulcer. Enterobacteriaceae, Butyricicoccus, Candidatus Soleaferrea, Lachnospiraceae NC2004 group, Peptococcus, and Enterobacteriales are considered to have a protective effect, indicating a negative correlation with the occurrence of Gastroduodenal ulcer. Conversely, Streptococcaceae, Lachnospiraceae UCG010, Marvinbryantia, Roseburia, Streptococcus, Mollicutes RF9, and NB1n are associated with a non-protective effect, demonstrating a positive correlation with Gastroduodenal ulcer incidence.
The scatter plot also displays the weights obtained through various MR analysis methods (IVW, MR-Egger, weighted median, weighted mode, and simple mode). These lines represent the non-protective or protective relationships between diverse gut microbiota and Gastroduodenal ulcer. An upward trend from left to right suggests a non-protective relationship with Gastroduodenal ulcer, while a downward trend indicates a protective relationship with Gastroduodenal ulcer (Supplementary Figures S3).
Through leave-one-out analysis, we identified no potential outliers among all instrumental variables (IVs), signifying that the established causal relationships remain unaffected by individual instrumental variables (IVs) (Supplementary Figures S1, S2). This finding further bolsters the reliability of the association between gut microbiota and Gastroduodenal ulcer.
4 DiscussionTo the best of our knowledge, this study represents the inaugural exploration into the causal association between gut microbiota and Gastroduodenal ulcer, utilizing publicly available GWAS data. Employing two-sample MR methods, we have effectively uncovered 13 causal relationships connecting gut microbiota to Gastroduodenal ulcer risk. This discovery furnishes pivotal scientific substantiation, advancing our comprehension of the impact of gut microbiota on Gastroduodenal ulcer etiology. These findings underscore the integral role played by the gut microbiota composition in Gastroduodenal ulcer. Further investigations hold the potential to deepen our insights into how these microbial entities influence the onset and progression of Gastroduodenal ulcer. Moreover, these revelations proffer novel perspectives for Gastroduodenal ulcer prevention and treatment strategies, including the prospect of modulating gut microbiota composition to enhance gastrointestinal well-being and curtail Gastroduodenal ulcer risk.
In our study, a series of analyses have indicated that a high abundance of Enterobacteriaceae may confer protection against Gastroduodenal ulcer. Previous research has demonstrated that Enterobacteriaceae exhibits resilience in acidic environments by inducing a low pH-triggered lysine decarboxylase system (CadB-CadA system). This mechanism converts lysine into cadaverine, an alkaline amine, which is subsequently released from the cells via CadB, leading to a reduction in extracellular hydrogen ion concentration (McGowan et al., 1996). The reduction in hydrogen ion concentration plays a significant role in effectively controlling the occurrence of Gastroduodenal ulcer. This discovery enhances our understanding of the protective capacity of Enterobacteriaceae against Gastroduodenal ulcer. Further investigation into the mechanisms underlying the protective role of Enterobacteriaceae in Gastroduodenal ulcer can pave the way for the development of more effective treatments. For instance, by intervening in the CadB-CadA system, we can potentially augment the survival capabilities of Enterobacteriaceae, thereby strengthening its protective effects against Gastroduodenal ulcer. Such interventions could have implications for the prevention and treatment of these ulcerative conditions. It’s worth noting that the gut’s indigenous microbial population includes Proteobacteria, a major constituent of the gut microbiota (Kim et al., 2017). In a study investigating alterations in the gut microbial community following a 14-day bismuth quadruple therapy for peptic ulcers, substantial changes were observed in the gut microbiota at the phylum level after the treatment period. There was a notable reduction in the abundance of specific gut bacteria at the phylum level. However, it is noteworthy that the abundance of Proteobacteria, which includes Enterobacteriaceae, exhibited a relative increase after the treatment (Zhou et al., 2020). The observed increase in Proteobacteria abundance implies a potential protective effect against Gastroduodenal ulcer. Notably, our study revealed that Enterobacterales, a taxonomic order within the Proteobacteria phylum, is associated with this increase, suggesting that a higher abundance of Enterobacterales may correspond to a reduced incidence of Gastroduodenal ulcer. Furthermore, it is essential to underline that Enterobacteriaceae, which is a family within Proteobacteria, aligns with these findings, further corroborating our research results.
In a study evaluating the efficacy of Helicobacter pylori eradication therapy for Gastroduodenal ulcer induced by this bacterium, researchers focused on a group of patients diagnosed with Helicobacter pylori infection who had not undergone any prior treatment. Within this cohort, Butyricicoccus was detected in the gut microbiota. Subsequent to treatment, a notable reduction in the abundance of Butyricicoccus was observed in comparison to both the uninfected Helicobacter pylori group and the control group devoid of severe digestive system ailments. Interestingly, the study identified a substantial increase in the abundance of Butyricicoccus among patients with Gastroduodenal ulcer before treatment in contrast to after treatment (Cui et al., 2022). This implies that an elevated abundance of Butyricicoccus may play a role in the development of Gastroduodenal ulcer, which contrasts with our findings. To reconcile this inconsistency, a more comprehensive understanding of the underlying mechanisms and principles is required to better elucidate the impact of Butyricicoccus on Gastroduodenal ulcer.
Xia Chen et al. conducted a study indicating a notably higher abundance of Streptococcus in patients with gastroduodenal ulcer. This observation implies that an elevated abundance of Streptococcus might be a risk factor for gastroduodenal ulcer, aligning with our own findings (Chen X. et al., 2018). It’s important to highlight that prior studies have detected Mollicutes in patients with chronic gastritis, but its abundance is comparatively lower in healthy individuals (Nascimento et al., 2021). This indicates that Mollicutes may have an impact on the occurrence of Gastroduodenal ulcer. Both RF9 and NB1n are categorized under Mollicutes, and our findings align with the notion that RF9 and NB1n may also exhibit a positive correlation with Gastroduodenal ulcer.
Candidatus Soleaferrea belongs to the Candida genus. In an experiment involving rats induced with cysteamine to induce Gastroduodenal ulcer perforation, the group administered with Candida exhibited a significantly higher probability of Gastroduodenal ulcer perforation compared to the group administered with normal saline. Furthermore, the area of Gastroduodenal ulcer was also larger in the Candida-administered group than in the normal saline group. These findings from the cysteamine-induced Gastroduodenal ulcer experiment indicate that Candida can significantly exacerbate Gastroduodenal ulcer (Nakamura et al., 2007). Additionally, there have been studies indicating that Candida infection is present in some patients with gastric-duodenal ulcers. Moreover, in cases where patients have both Gastroduodenal ulcer and Barrett’s ulcers, Candida is observed exclusively in those with Gastroduodenal ulcer (Kalogeropoulos and Whitehead, 1988). Nonetheless, our experimental results revealing a negative correlation between increased Candidatus Soleaferrea abundance and Gastroduodenal ulcer contradict our initial hypothesis. This suggests the presence of other factors or mechanisms that may influence the relationship between Candidatus Soleaferrea and Gastroduodenal ulcer. Therefore, further research is warranted to comprehensively comprehend the association between Candidatus Soleaferrea and Gastroduodenal ulcer. This may involve investigating other potential microbiota alterations, host genetic factors, environmental influences, and more to elucidate the specific role of Candidatus Soleaferrea in Gastroduodenal ulcer occurrence. The findings from these studies will contribute to a deeper understanding of the interaction between Candidatus Soleaferrea and Gastroduodenal ulcer, offering fresh insights into potential treatment strategies and preventive measures.
Streptococcaceae, Lachnospiraceae NC2004 group, Lachnospiraceae U-CG010, Marvinbryantia, Peptococcus, and Roseburia all fall within the Firmicutes phylum. Firmicutes is a prevalent bacterial phylum typically identified in the human gut. The gut microbiota forms a multifaceted ecosystem comprising diverse microorganisms that exert significant influences on human health and disease (Belkaid and Hand, 2014; Sender et al., 2016; Thursby and Juge, 2017). Recent studies have indicated that patients infected with Helicobacter pylori tend to exhibit higher Firmicutes abundance in their gut microbiota prior to treatment. However, following a 14-day course of bismuth therapy, significant alterations occur within the gut microbial community, marked by a substantial reduction in Firmicutes abundance (Chen L. et al., 2018). In a study involving mice with Helicobacter pylori-induced gastritis, researchers observed an elevated abundance of Firmicutes. These findings indicate a potential positive correlation between increased Firmicutes abundance and the risk of Gastroduodenal ulcer (Lofgren et al., 2011). In our study, we identified a positive correlation between the abundance of Lachnospiraceae UCG010, Marvinbryantia, Streptococcaceae, and Roseburia and the incidence of Gastroduodenal ulcer. This suggests that an increase in the abundance of these bacteria may be associated with a higher risk of Gastroduodenal ulcer. However, the specific relationship between Firmicutes and Gastroduodenal ulcer remains unclear due to limited research in this area. The scientific community has yet to establish a consensus on this matter, necessitating further investigation to confirm these findings.Conversely, our research revealed a negative correlation between the abundance of Lachnospiraceae NC2004 group, and Peptococcus and Gastroduodenal ulcer. However, due to the scarcity of relevant studies, we do not have a comprehensive understanding of the specific relationship between these bacterial genera and Gastroduodenal ulcer. Thus, additional research is warranted to elucidate the associations between these genera and the occurrence and progression of Gastroduodenal ulcer. Through in-depth investigations and experiments, we can gain a better understanding of how these bacterial genera contribute to the development of Gastroduodenal ulcer.
Our study employed Mendelian Randomization (MR) analysis methods, which, in comparison to traditional observational studies, can mitigate the influence of confounding factors on the outcomes. We conducted an assessment of the causal relationship between gut microbiota and Gastroduodenal ulcer, specifically at the phylum level. This analysis serves as a foundational framework for future investigations into specific microbial strains, thereby contributing to a deeper comprehension of the pathogenesis of Gastroduodenal ulcer. Our study’s findings offer new insights and potential approaches for the future diagnosis and treatment of Gastroduodenal ulcer. Simultaneously, in order to assess the potential impact of heterogeneity and pleiotropy among instrumental variables on the MR results, we conducted an extensive sensitivity analysis, which further bolsters the reliability of our findings.
Our study has certain limitations that should be taken into account. Firstly, the participants included in the GWAS meta-analysis database were predominantly of European descent, with a limited amount of data from other ethnic groups regarding their gut microbiota. This discrepancy may have influenced our research results, as the composition of gut microbiota can vary among different ethnic groups. And, due to the fact that most of the GWAS data comes from individuals of European descent, even if there may be interference from population stratification, the results of this study may not be applicable to other populations of non-European ancestry. Secondly, in our study, 16S rRNA gene sequencing enables resolution only at the phylum level, preventing us from further exploring the causal relationship between gut microbiota and gastric ulcers at the species level. Moreover, concerning sample size, gut microbiota Genome-Wide Association Studies (GWAS) are in an early stage, with relatively few loci associated with gastric ulcers. To conduct sensitivity analysis and test at various significance levels, more genetic variations need inclusion as instrumental variables. Further exploration requires analysis at higher taxonomic levels, such as order, class, and phylum, which might limit the comprehensive study of specific impacts of individual bacterial species. Thirdly, using a limited number of gut microbiota Single Nucleotide Polymorphisms (SNPs) as instrumental variables, there’s a possibility that our study results could be influenced by weak instrument bias, despite all genetic instruments being associated with the exposure (F-statistic>10). It’s noteworthy that our study predominantly involved individuals of European descent, potentially limiting the generalizability of our study results to a more diverse population.
Furthermore, it’s essential to acknowledge that MR analysis is a hypothesis-based approach, and its outcomes can only establish associations rather than causal relationships. Subsequent experimental and clinical research is indispensable to establish the causal relationship between gut microbiota and specific diseases.
Additionally, there might be some subjectivity involved in eliminating the confounding effects of genetic variables through phenoscanner. This subjectivity could introduce some bias into our research results, emphasizing the importance of interpreting and understanding the findings with caution.
In conclusion, although our study has uncovered valuable insights, it’s imperative to recognize the aforementioned limitations. Future research should aim for more comprehensive and diversified investigations to further enhance our understanding of the intricate relationship between gut microbiota and diseases.
In summary, our study conducted a comprehensive assessment of the causal relationship between gut microbiota and Gastroduodenal ulcer. Our findings have contributed valuable insights and directions for further research on the prevention and treatment of Gastroduodenal ulcer. However, while we have acquired an initial understanding of the connection between gut microbiota and Gastroduodenal ulcer, the precise mechanisms underlying the role of gut microbiota in this condition remain unclear. Our study has established a correlation, but further research is necessary to elucidate how gut microbiota influences the occurrence and progression of Gastroduodenal ulcer.
In future investigations, we plan to delve deeper into the mechanisms through which gut microbiota contributes to the development of Gastroduodenal ulcer. This will involve analyzing the composition and functionality of the gut microbiota and its interactions with the host. Such endeavors will enhance our comprehension of the relationship between gut microbiota and Gastroduodenal ulcer, offering more targeted approaches for the prevention and treatment of this condition.
Moreover, while our study has uncovered pivotal insights into the relationship between gut microbiota and Gastroduodenal ulcer through Mendelian Randomization (MR), it’s essential to acknowledge the inherent limitations of this method. MR, by nature, establishes associations but doesn’t conclusively prove causation. It relies on genetic variance, lacking the capacity for direct exposure manipulation present in randomized controlled trials. Hence, it’s imperative to recognize the need for additional experimental and clinical research to validate and establish causative relationships between specific exposures and health outcomes, which can provide more direct and conclusive evidence.
Data availability statementThe datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributionsYW: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Supervision, Writing – original draft, Writing – review & editing. JZ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. YH: Conceptualization, Data curation, Investigation, Methodology, Software, Supervision, Writing – original draft, Writing – review & editing. LW: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Project administration, Software, Writing – original draft, Writing – review & editing. QZ: Conceptualization, Data curation, Formal analysis, Methodology, Supervision, Writing – original draft. BH: Formal Analysis, Project administration, Resources, Visualization, Writing – review & editing. ZL: Formal analysis, Funding acquisition, Project administration, Resources, Validation, Visualization, Writing – original draft.
FundingThe author(s) declare financial support was received for the research, authorship, and/or publication of this article. The current study was supported by Science and Technology Research Project of Jiangxi Education Department (GJJ2200116; GJJ2200172); Science and Technology Research Project of Jiangxi Provincial Administration of Traditional Chinese Medicine (2020A0130; 2022A342); Science and Technology Project of Jiangxi Health Committee (Project Number: 20161999; 20203330; 202310482).
AcknowledgmentsWe want to acknowledge the participants and investigators of the FinnGen study and the MiBioGen consortium.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1322537/full#supplementary-material
ReferencesArnold, M., Raffler, J., Pfeufer, A., Suhre, K., Kastenmuller, G. (2015). Snipa: an interactive, genetic variant-centered annotation browser. Bioinformatics 31 (8), 1334–1336. doi: 10.1093/bioinformatics/btu779
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowden, J., Davey, S. G., Haycock, P. C., Burgess, S. (2016a). Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet. Epidemiol. 40 (4), 304–314. doi: 10.1002/gepi.21965
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowden, J., Del, G. M. F., Minelli, C., Davey, S. G., Sheehan, N. A., Thompson, J. R. (2016b). Assessing the suitability of summary data for two-sample mendelian randomization analyses using mr-egger regression: the role of the i2 statistic. Int. J. Epidemiol. 45 (6), 1961–1974. doi: 10.1093/ije/dyw220
PubMed Abstract | CrossRef Full Text | Google Scholar
Burgess, S., Butterworth, A., Thompson, S. G. (2013). Mendelian randomization analysis with multiple genetic variants using summarized data. Genet. Epidemiol. 37 (7), 658–665. doi: 10.1002/gepi.21758
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, L., Xu, W., Lee, A., He, J., Huang, B., Zheng, W., et al. (2018). The impact of helicobacter pylori infection, eradication therapy and probiotic supplementation on gut microenvironment homeostasis: an open-label, randomized clinical trial. Ebiomedicine 35, 87–96. doi: 10.1016/j.ebiom.2018.08.028
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, X., Xia, C., Li, Q., Jin, L., Zheng, L., Wu, Z. (2018). Comparisons between bacterial communities in mucosa in patients with gastric antrum ulcer and a duodenal ulcer. Front. Cell Infect. Microbiol. 8. doi: 10.3389/fcimb.2018.00126
CrossRef Full Text | Google Scholar
Cui, M. Y., Cui, Z. Y., Zhao, M. Q., Zhang, M. J., Jiang, Q. L., Wang, J. J., et al. (2022). The impact of helicobacter pylori infection and eradication therapy containing minocycline and metronidazole on intestinal microbiota. BMC Microbiol. 22 (1), 321. doi: 10.1186/s12866-022-02732-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Hartwig, F. P., Davey, S. G., Bowden, J. (2017). Robust inference in summary data mendelian randomization via the zero modal pleiotropy assumption. Int. J. Epidemiol. 46 (6), 1985–1998. doi: 10.1093/ije/dyx102
PubMed Abstract | CrossRef Full Text | Google Scholar
Hemani, G., Tilling, K., Davey, S. G. (2017). Orienting the causal relationship between imprecisely measured traits using gwas summary data. PloS Genet. 13 (11), e1007081. doi: 10.1371/journal.pgen.1007081
PubMed Abstract | CrossRef Full Text | Google Scholar
Holmes, M. V., Ala-Korpela, M., Smith, G. D. (2017). Mendelian randomization in cardiometabolic disease: challenges in evaluating causality. Nat. Rev. Cardiol. 14 (10), 577–590. doi: 10.1038/nrcardio.2017.78
PubMed Abstract | CrossRef Full Text | Google Scholar
Huang, J. Q., Sridhar, S., Hunt, R. H. (2002). Role of helicobacter pylori infection and non-steroidal anti-inflammatory drugs in peptic-ulcer disease: a meta-analysis. Lancet 359 (9300), 14–22. doi: 10.1016/S0140-6736(02)07273-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Kalogeropoulos, N. K., Whitehead, R. (1988). Campylobacter-like organisms and candida in peptic ulcers and similar lesions of the upper gastrointestinal tract: a study of 247 cases. J. Clin. Pathol. 41 (10), 1093–1098. doi: 10.1136/jcp.41.10.1093
PubMed Abstract | CrossRef Full Text | Google Scholar
Kamat, M. A., Blackshaw, J. A., Young, R., Surendran, P., Burgess, S., Danesh, J., et al. (2019). Phenoscanner v2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics 35 (22), 4851–4853. doi: 10.1093/bioinformatics/btz469
PubMed Abstract | CrossRef Full Text | Google Scholar
Kim, S., Covington, A., Pamer, E. G. (2017). The intestinal microbiota: antibiotics, colonization resistance, and enteric pathogens. Immunol. Rev. 279 (1), 90–105. doi: 10.1111/imr.12563
PubMed Abstract | CrossRef Full Text | Google Scholar
Kurilshikov, A., Medina-Gomez, C., Bacigalupe, R., Radjabzadeh, D., Wang, J., Demirkan, A., et al. (2021). Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 53 (2), 156–165. doi: 10.1038/s41588-020-00763-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Kurki, M. I., Karjalainen, J., Palta, P., Sipila, T. P., Kristiansson, K., Donner, K. M., et al. (2023). Finngen provides genetic insights from a well-phenotyped isolated population. Nature 613 (7944), 508–518. doi: 10.1038/s41586-022-05473-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Lofgren, J. L., Whary, M. T., Ge, Z., Muthupalani, S., Taylor, N. S., Mobley, M., et al. (2011). Lack of commensal flora in helicobacter pylori-infected ins-gas mice reduces gastritis and delays intraepithelial neoplasia. Gastroenterology 140 (1), 210–220. doi: 10.1053/j.gastro.2010.09.048
PubMed Abstract | CrossRef Full Text | Google Scholar
McGowan, C. C., Cover, T. L., Blaser, M. J. (1996). Helicobacter pylori and gastric acid: biological and therapeutic implications. Gastroenterology 110 (3), 926–938. doi: 10.1053/gast.1996.v110.pm8608904
PubMed Abstract | CrossRef Full Text | Google Scholar
Miftahussurur, M., Yamaoka, Y. (2015). Helicobacter pylori virulence genes and host genetic polymorphisms as risk factors for peptic ulcer disease. Expert Rev. Gastroenterol. Hepatol. 9 (12), 1535–1547. doi: 10.1586/17474124.2015.1095089
PubMed Abstract | CrossRef Full Text | Google Scholar
Milne, R. L., Kuchenbaecker, K. B., Michailidou, K., Beesley, J., Kar, S., Lindstrom, S., et al. (2017). Identification of ten variants associated with risk of estrogen-receptor-negative breast cancer. Nat. Genet. 49 (12), 1767–1778. doi: 10.1038/ng.3785
PubMed Abstract | CrossRef Full Text | Google Scholar
Nakamura, T., Yoshida, M., Ishikawa, H., Kameyama, K., Wakabayashi, G., Otani, Y., et al. (2007). Candida albicans aggravates duodenal ulcer perforation induced by administration of cysteamine in rats. J. Gastroenterol. Hepatol. 22 (5), 749–756. doi: 10.1111/j.1440-1746.2006.04353.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nascimento, A. C., Amorim, A. T., Barbosa, M. S., Alexandre, J., Campos, G. B., Macedo, C. L., et al. (2021). Evaluating the presence of mycoplasma hyorhinis, fusobacterium nucleatum, and helicobacter pylori in biopsies of patients with gastric cancer. Infect. Agent. Cancer. 16 (1), 70. doi: 10.1186/s13027-021-00410-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Schulz, C., Schutte, K., Koch, N., Vilchez-Vargas, R., Wos-Oxley, M. L., Oxley, A., et al. (2018). The active bacterial assemblages of the upper gi tract in individuals with and without helicobacter infection. Gut 67 (2), 216–225. doi: 10.1136/gutjnl-2016-312904
PubMed Abstract | CrossRef Full Text | Google Scholar
Staley, J. R., Blackshaw, J., Kamat, M. A., Ellis, S., Surendran, P., Sun, B. B., et al. (2016). Phenoscanner: a database of human genotype-phenotype associations. Bioinformatics 32 (20), 3207–3209. doi: 10.1093/bioinformatics/btw373
留言 (0)