Protein tyrosine phosphatases are named based on their amino acid substrate that is believed to be involved in the catalytic transfer of phosphate group. Protein tyrosine phosphatase 1B (PTP1B) is well established as having association with negative regulation of insulin and progression of type II diabetes [1]. Recently, research has proven that PTP1B is also linked with inhibition of Alzheimer's disease related disorders [2]. Phosphorylation is very important for the regulation of signal transduction pathways and if disrupted, major human diseases are caused that include obesity and diabetes [[3], [4], [5]]. PTP1B is involved in the removal of phosphate group through catalysis. The transfer of phosphate group occurs in a cycle of reactions that results in open and closed conformations of protein.
There are 435 residues in PTP1B enzyme having a conserved domain at the N-terminus known as the PTP domain. There is a signature motif of (H/V)CXXXXXR(S/T) present in all the members of PTP superfamily [6]. In catalysis, critical residues are cysteine and arginine from the conserved PTP domain. When the ligand comes and binds to the enzyme, it binds in the phosphate binding site resulting in the closed conformation of a WPD loop. Amino acids forming the WPD loop are: TRP179-PRO180-ASP181. The chain is in open conformation when there is no ligand present at the catalytic site. There is also a phospho-intermediate form when phosphate group is present at the catalytic site and bound to cysteine residue resulting in cysteine-phosphate. When the ligand occupies the active pocket, the loop makes a move of 8 Å towards the pocket, “closing” it [6].
A common catalytic mechanism is observed among PTP family that has two main steps. In the first step, a phosphotyrosine substrate is involved that phosphorylates a cysteine residue and itself gets dephosphorylated. A thiophosphoryl bond is formed with the phosphate group of the substrate as the side chain acts as a nucleophile. Arginine residue that belongs to the PTP characteristic motif plays its role in substrate binding and hence stabilizes the transition state. WPD loop also has no lesser role, the deprotonated carboxylate group of aspartic acid plays a part of providing a base during cleavage of phosphate group in the phospho-intermediate form. These catalytic processes can help in hydrolysis with the placement of water molecule at correct binding site of the enzyme [6,7].
Metal ions perform critical structural and functional roles in biochemical processes such as metabolic regulation, energy conversion, material transportation, and information transmission in our body [8]. Hence, researchers are paying considerable attention to explore the clinical use of metal-based complexes in treating various ailments. Among these, Diabetes mellitus (DM) is a chronic illness that affects a vast majority of population globally. Presently used synthetic drugs for diabetes have several side effects including mental and physical pain due to daily insulin injections [9]. Therefore, the concept of treating diabetes using metal complexes such as zinc has been introduced as an alternative to commonly used therapeutics. In a study by Fugono and coworkers, in vitro studies were performed on zinc(II) complexes to investigate their insulinomimetic activity in isolated rat adipocytes. In addition to this, in vivo hypoglycemic potential of complexes was also explored in type 2 diabetic model KK-Ay mice through daily intraperitoneal injections. Findings of the study concluded that zinc(II) complexes have the potential to treat diabetes when orally administered in rat model [10]. Following this, more in vivo studies were also conducted using different zinc(II) complexes to explore the antidiabetic potential of metallo-complex in controlling induced experimental diabetes in rats. The results depicted that the antidiabetic potential of investigated zinc(II) complex was comparable with the standard antidiabetic drug gliclazide [11]. Along with the experimental, advanced computational studies are also carried out to gain insights into the zinc ion inhibition of PTP1B while highlighting the role of zinc ion as insulin-mimetic [12]. The mechanism by which zinc ion acts as insulin-mimetic has not yet been fully explored but significant amount of research evidence points towards the role of PTP1B. Inhibition of PTP1B activates insulin receptors which is associated with the activation of cytosolic nonreceptor tyrosine kinase followed by sequence of events which finally lead to the translocation of glucose transporter 4 (GLUT4) and activation of phosphodiesterase [13].
Although computational and experimental studies have revealed that zinc ion has insulin-mimetic activity, the interaction mechanism and structural dynamics of zinc(II) complexes is not explored till yet [14]. Computational chemistry is a valuable tool to investigate metal-ion containing systems. However, modeling metal ions complexes to probe their behavioral dynamics is arduous in biological systems. Zinc AMBER force field (ZAFF) has been developed by Peters and coworkers which provides parameterized zinc metal centers that can be straightforwardly used in tleap [15]. However, in our case, zinc metal center (Zn-OOOO) that needs to be simulated did not exist in ZAFF. Therefore, an ab initio metal ion modeling using a python based metal center parameter builder (MCPB.py) was performed which employs the bonded model approach [16]. Based on the reported observation that zinc ions inhibit PTP1B, the objective of the present investigation was to identify zinc containing ligands as inhibitors for PTP1B. Comparative dynamics studies were performed on three PTP1B 3D structures (PDB id: 2CM2, 3I80, and 1A5Y) to elucidate the binding site and binding of metal complexes inside the pocket of different conformational states of the enzyme.


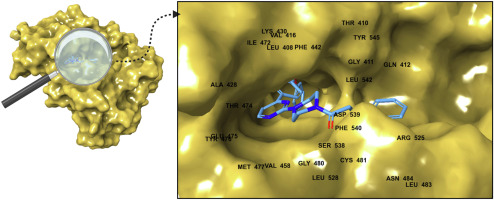
Comments (0)